中国科研团队揭示暹罗斗鱼表型多样性的遗传秘密

#头条创作挑战赛#
↑↑↑关注我,医学,药物,健康,科研,每天一篇世界最前沿原创文章。
暹罗斗鱼(Betta splendens)原产于泰国中部和湄公河下游,以其驯化形式而闻名,被认为是观赏鱼,但最初是为了在类似于斗蟋蟀的赌博比赛中使用而繁殖的。通过圈养繁殖,出现了各种各样的行为和形态,包括侵略性,色素沉着,体型和鳍形状的变化。

色彩斑斓的暹罗斗鱼
对科研和养宠物鱼感兴趣的小伙伴点击链接看看:斑马鱼的消毒方法,也适用于金鱼等宠物鱼~~
暹罗斗鱼易于繁殖和养殖,并且由于具有高度的种内变异性和大量表征表型,为探索脊椎动物行为和形态的遗传基础提供了有用的资源。它还展示了一个人类的直接和间接选择如何塑造了一个驯化的物种的典型例子。
近日,南昌大学生命科学学院洪一江教授团队报告了雌性双歧杆菌的高效染色体级基因组组装,对来自暹罗斗鱼复合体中其他六个物种的727个驯化个体和59个野生个体的重测序数据。研究人员研究了品种之间的进化关系和起源,并使用关联映射来识别许多不同特征的遗传基础,包括SD,鳍形态,着色,体型和侵略性以及其他行为。
暹罗斗鱼的基因组组装、注释和比较分析
研究人员使用多方面的测序和组装工作流程生成了贝塔鱼的高质量染色体级组装,包括PacBio读数,Illumina读数,Hi-C读数,10x基因组学读数和BioNano光学映射。最终组装的基因组为4.5129亿个碱基对,其中连续体和支架N50分别达到4.07和19.63 Mb。共有93.6%的支架被放置在21条染色体上,这与之前报道的染色体核型一致。
该组件包含 119 Mb 的重复序列和25,104个注释的结构基因,其中22,788个(90.77%)是功能注释的。核心真核基因图谱方法分析证实,248个(96.4%)完整核心真核基因中存在239个,基准通用单拷贝直系同源物评估显示,2586个(97.5%)单拷贝同源基因中有2522个被注释,表明基因组和基因注释的完整性高。
研究人员使用由暹罗斗鱼和其他13个硬骨质共享的465个单拷贝同源物构建了一个系统发育,并表明暹罗斗鱼在大约1.096亿年前与其他鲈形目鱼分化。高质量的基因组组装加上全面的基因组注释,是对暹罗斗鱼研究现有参考基因组资源的重要补充和改进。
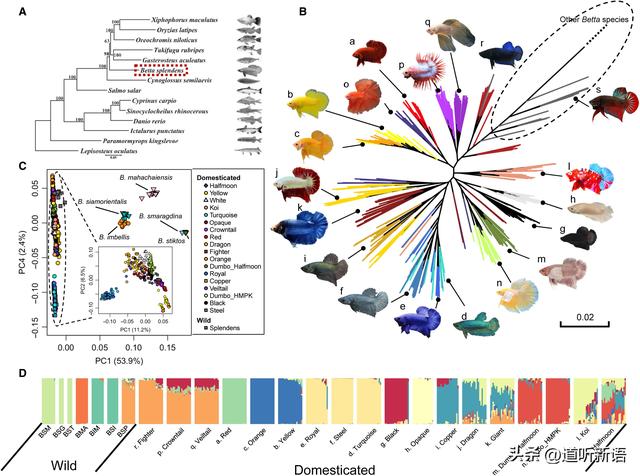
(A)硬骨鱼的系统发育(B)串联全基因组单核苷酸多态性 (C) 暹罗斗鱼的PCA。(D)对双歧杆菌的混合分析。
贝塔鱼在驯化过程中的多样化
研究人员收集了14个品种的暹罗斗鱼,它们在尾巴类型,颜色,性别和体型方面各不相同并对727个个体进行了全基因组重测序,产生了~2.7 Tb的测序数据。
为了阐明暹罗斗鱼复合体的种群历史和驯化的暹罗斗鱼的起源,研究者进一步对来自暹罗斗鱼复合体的六个野生物种的59个个体进行了测序,对每个品种的20个随机挑选的个体进行共分析。来自串联序列的最大似然系统发育,代表平均基因组聚结树,表明驯化的品种相对于其他野生物种形成单系群。此外,在主成分分析中,它们始终如一地形成一个紧密的簇,与其他野生个体不同。
这些观察结果与以下假设相一致,即暹罗斗鱼的所有当前品种都是从同一组野生双歧杆菌中驯化的。贝塔西亚莫林塔个体在树中的贝塔英伯利斯个体簇中形成单系分支。斗鱼品种的几个分支作为外群体落在系统发育树中其他驯化品种的外群体,这与育种记录相符,即斗士品种代表了早期的驯化形式,最早的驯化贝塔鱼实际上是为战斗而选择的品种。
然而,这些观察结果也可以归因于野生物种对斗鱼,面尾和冠尾品种的渗透,因为在不同的鱼类群体中观察到广泛的基因流动信号。种群结构,如系统发育,PCA,和外加剂分析,表明由颜色和形态定义的品种通常聚集在一起。
尽管研究者注意到这一结论可能受到此处使用的采样策略的影响。半月普拉卡特品种是短鳍类型,与战斗型和野生暹罗斗鱼不同,具有圆形的尾巴形状和体形,也根据外观进行集群,主要与颜色有关。红色,黄色,橙色,绿松石绿,皇家蓝色,钢蓝色的品种组具有大量的群体特异性遗传漂移,这在结构分析和全基因组系统发育中是显着的,表明这些群体在其驯化史上经历了强烈的瓶颈效应。
混合分析表明暹罗斗鱼野生和驯化物种之间存在大量基因流动。这些结果显示出驯化物种和野生物种之间基因流动量的普遍异质性,甚至在同一品种的个体中也很明显。个体间混合物的高度异质性表明,基因流动不仅在野生和驯化的品种中很常见,而且是最近才出现的,可能是由于人为的影响。至少有两种野生物种,贝塔马哈柴氏鱼和双歧杆菌,为驯养的暹罗斗鱼的基因组组成做出了贡献。因此也可能促成了这些品种的表型变异。
dmrt1中的调节变体介导了双歧杆菌的雄性异配子
为了绘制位点,研究者使用混合线性模型对所有727个个体进行了关联映射,包括590名雄性和137名雌性。将亲缘矩阵和PCA的前三个主要成分用作协变量,以最大限度地减少种群分层的影响。
研究人员在9号染色体上的位置27.75至27.81 Mb处发现了一个非常重要的图谱位置。与不同品种的映射一致地重复信号,支持所有这些品种中SD的共同遗传基础,这也与以前的研究一致。在537个具有铅变异杂合基因型的个体的537个个体中,有533个个体是表型雄性,137个AA纯合子个体中有130个是雌性,这强烈支持了暹罗斗鱼雄性异配子(XY /XX)的假设。
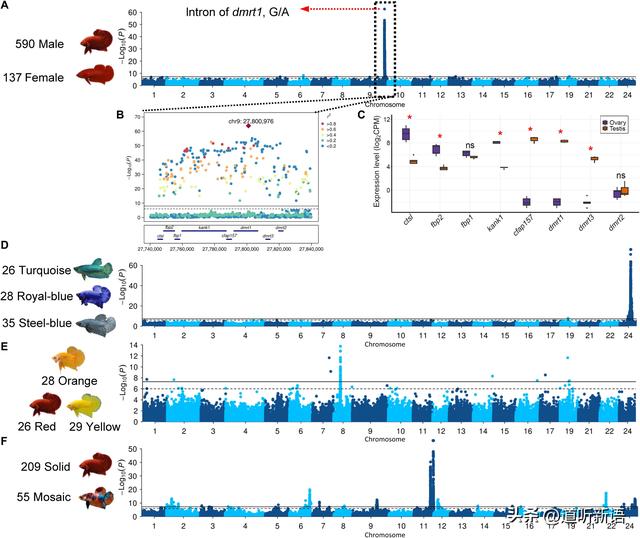
A)SD的GWAS分析 B)相关峰区上的分布图。 C)卵巢和睾丸基因的表达水平。D)表型的曼哈顿图 F)颜色图案表型
最强关联的变体簇以 dmrt1 和 kank1 的内含子组成。kank1通过调节肌动蛋白聚合在细胞骨架形成中起作用,这使得它不太可能成为SD。相比之下,dmrt1是一种众所周知的基因,有助于鱼类,鸟类和爬行动物中的SD。
此外,mRNA测序显示dmrt1在睾丸中高度表达,但在卵巢中几乎检测不到。暹罗斗鱼中的dmrt1在进化上接近青鳉鱼dmy基因,这是在硬骨质中鉴定出的第一个性别决定基因。同配子雌性基因型雄性的存在可能表明环境因素,如温度,也可能像许多其他鱼类一样在SD中发挥作用。
其他野生物种,包括双歧杆菌、西亚莫里塔利斯、马哈恰伊氏双歧杆菌、斯马拉吉纳双歧杆菌、吉他双歧杆菌和双歧杆菌,可能具有不同的SD机制,因为雌性和雄性个体在领先的单核苷酸多态性(SNP)处的G等位基因都是纯合子的,并且与这些个体的全基因组关联研究(GWAS)没有在dmrt1处恢复信号 位点。
贝塔鱼的颜色和颜色图案
研究表明,钢蓝色品种暹罗斗鱼与其他品种的不同之处在于鳞片下层缺乏红细胞。将钢蓝、皇家蓝和绿松石绿编码为 1、2和3的GWA分析在24号染色体上以 8.96 至 9.19 Mb 的速度识别出单个位点,以及具有其他编码方案的 GWAS 始终映射到同一位置。
SNP峰值的基因型分析表明,绿松石绿和钢蓝是不同等位基因的纯合子,28个皇家蓝个体中有27个是杂合子,这与共显性遗传模式高度一致。该位点包含13个蛋白质编码基因,包括mthfd1l。mthfd1l参与四氢叶酸的合成,四氢叶酸从事嘌呤的从头组装,嘌呤是产生虹彩团的关键成分,产生虹彩颜色,包括蓝色和绿色。
因此,mthfd1l是一个有前途的候选者,需要深入的功能研究。铜品种暹罗斗鱼的鳞片上具有特征性的金属外观。繁殖记录表明,它是通过将“金属基因”引入钢蓝品种的遗传背景而衍生出来的。
进行了GWAS,确定了两个最重要的峰。最强的一个与皇家蓝色,钢蓝色和绿松石绿色变化下面的位点重叠,另一个位于5号染色体的10.37 Mb处。所有峰变体都存在于srgap3基因中,细胞骨架调节剂。该位点的先导SNP的基因型分析显示,钢蓝是T等位基因的纯合子,而铜个体携带一个或两个替代等位基因(C)的拷贝,与已知的遗传模式一致。
因此,研究者认为相关区域是假设的孟德尔金属基因的有力候选者。在硬骨质中,红细胞团和黄色团分别产生红色和黄色颜料,橙色可以由蝶啶组分或红色和黄色颜料的混合物赋予。案例对照GWAS对比橙色品种与红色 和黄色品种在8号染色体5.83 Mb的位置鉴定出一个主要位点,包含93个变体,全部位于rnf213基因中,参与血管生成和血管发育中的非规范Wnt信号通路。
红色和黄色品种是T等位基因缺失的纯合子,而橙色品种的TA等位基因插入频率较高。马赛克彩色花纹是暹罗斗鱼中一种壮观的表型,具有多种商业名称,包括锦鲤,糖果,星系,柠檬和大理石。虽然在锦鲤和青鳉鱼中也观察到马赛克颜色图案,但尚未研究潜在的分子基础。
在这里,研究者执行了GWAS,在实心和马赛克颜色 之间进行案例控制设计。并在八条染色体上鉴定出九个相关位点,提示该表型的多基因基础。两个最强的信号在11号染色体上的两个相邻峰中发现。在最重要的位点上,重点介绍与色素沉着有关的slc39a7,chs3,chs8和col11a2,以及参与细胞内色素动员的管状物。此外,相邻的轨迹还含有色素沉着相关基因,包括臀位素,eppk1,slc17a5和slc52a2。
值得注意的是,在关联峰中,有八个拷贝的plec,这是一个基因共表达网络分析中的中心基因,用于洞穴鱼中“粉红色 - 深绿色”颜色变化。暹罗斗鱼的眼睛颜色也表现出极大的多样性,至少六种颜色类别,包括黑色,白色,黄褐色,黄色,浅蓝色和棕色,可以视觉识别。然而,每个组的GWA映射并未显示强相关位点,可能表明眼睛颜色变化背后的复杂遗传结构或环境线索。
kcnj15的功能增益导致鳍片过度生长
暹罗斗鱼之间最显着的形态差异在于鳍,特别是尾鳍。面纱尾 和半月在背鳍,肛鳍和尾鳍方面显示出与战斗型相比的显着增长。杂交种表明所有长鳍品种具有共同的遗传基础,长鳍表型优于短鳍表型。
为了绘制潜在的遗传变异,进行了GWAS对比长鳍与短鳍个体。一个极显著位点以14号染色体9.60 Mb为中心的位置被鉴定出来。完全链接中有三种导联变体:一种同义突变,一种位于smg8中的内旋,一个位于kcnj15的3'未翻译区域(3′UTR)。
所有三种变体都完美地区分了驯化个体中的长鳍和短鳍。smg8蛋白充当激酶活性的调节剂,参与无意义介导的mRNA衰变,并且不太可能成为长鳍表型的候选者。相比之下,kcnj15编码钾通道,这是一个更好的候选者,因为编码钾通道的几个基因,包括kcnk5b,kcnh2a,kcnj13和kcc4a,已被鉴定为在斑马鱼中引起各种长鳍表型。
此外,通过RNA测序(RNA-seq)比较长鳍和短鳍个体之间尾鳍中两个候选基因的表达谱时,发现smg8的表达水平在不同鱼类组之间相似,而kcnj15在长鳍品种中表达量高,显示出显着差异,其转录本在短鳍品种中几乎检测不到。
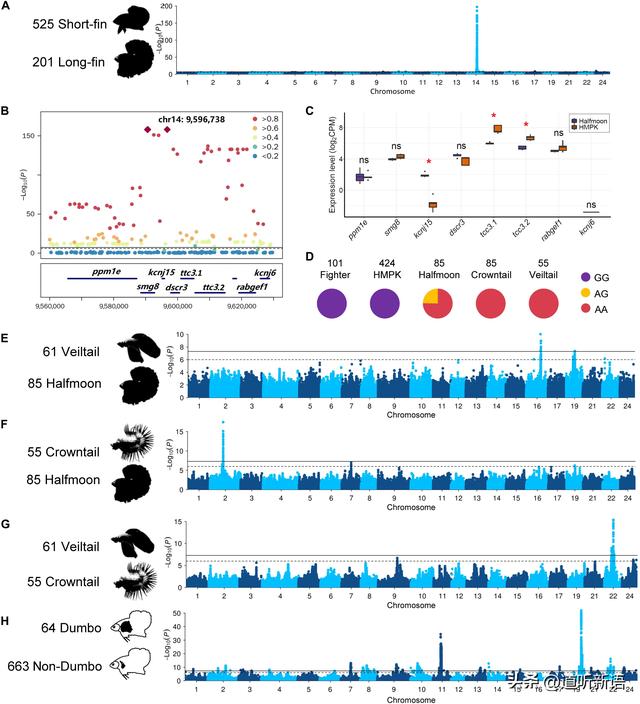
A)长鳍与短鳍形态图。B)全基因组信号图。C)尾鳍基因谱。 D)双歧杆菌SNP。 E to G)曼哈顿图, H)GWAS图。
大多数野生物种,包括灿烂双歧杆菌、双歧杆菌、马哈柴氏双歧杆菌和双歧杆菌,都被认为是短鳍,并且始终将所有这些个体固定在kcnj15的SNP处的G等位基因,并且很少能检测到转录本。在斯马拉格迪纳双歧杆菌和斯玛拉吉纳双歧杆菌吉他群体中,该变体仍在分离。在杂合子斯马拉吉纳双歧杆菌个体中,可以检测到预测的转录本,但表达水平较低,这可能是由于长尾品种最近在圈养繁殖过程中的入侵造成的。鉴于先导变体位于3′UTR上,并且变体等位基因与kcnj15的表达水平之间存在强关联,推测该变体可能影响了kcnj15的mRNA稳定性。
面尾、冠尾和半月表型
在长鳍品种中,还有很多额外的表型变异,面尾,冠尾和半月品种是一些最独特的品种。面尾鱼是记录的第一个长鳍品种,而后来选择尾鳍鳐完全180°传播产生了半月形品种。冠尾品种与半月品种的不同之处在于鳍鳍之间的织带组织减少。
GWAS对比面纱尾和半月在16号染色体上鉴定出13.48至13.79 Mb的300 kb区域,可能与鳍扩散表型有关。在该位点中,有四个基因(znf407,zadh2,tshz1和znf516)编码锌指蛋白,广泛参与转录调节和细胞功能。GWAS比较半月和冠尾在2号染色体上显示12.21至12.27 Mb的显着信号,可能与织带表型有关。
在位点内,一簇显着相关的变异位于a0zsk3(新膀胱毒素亚基α)和cep70(中心体蛋白70)之间的基因间区域,其没有任何与表型直接相关的功能,可能表明潜在因果变异的调节作用。比较冠尾和面尾品种,发现22号染色体上有一个位点(8.25至8.40 Mb)跨越八个基因,包括 frmd6,它参与肌动蛋白结构组织,并且具有与上皮到间充质过渡特征相关的表达丧失,使其成为解释冠尾和面尾品种之间鳍形态表型差异的强候选基因。
“小飞象”表型
硬骨动物的胸鳍与两栖动物,爬行动物和哺乳动物的前附件同源。一个品种,小飞象,其特征是其成对的胸鳍过度生长,具有更多和细长的鳍射线。使用小飞象作为案例和非小飞象作为对照的GWAS识别出两个强信号,它们分别位于11号 和 19染色体上。
在基因表达分析的基础上,因果基因可能是kcnh8。然而,该基因位于我们研究中确定的11号染色体上的相关位点之外。此外,在比较小飞象和非小飞象品种时,在胸鳍组织中没有检测到kcnh8的差异表达。
研究中的主要SNP位于一个包含来自hoxa基因家族的八个基因簇的区域,这对于形成硬骨质中的鳍骨骼和手指至关重要。对于19号染色体上的信号,其中一个SNP位于fbxl15的3′UTR上,这是一种参与背/腹侧模式形成和骨鲈维持的基因,使其成为一个强大的候选基因。
巨型表型
体型在许多生物体中表现出多基因遗传,包括人类。在暹罗斗鱼中,巨型突变体显示出显着的身体扩大,如总长度,标准长度,重量的增加。对照GWAS使用体长测量结果将大小与正常大小的品种和GWAS进行比较,在8号染色体的2.03至2.26 Mb位置发现了一个共同的重要位点,这解释了8.1%至9.0%的表型方差。
仔细检查位点中的基因,研究者没有发现编码全球身体发育调节因子的基因,这是预期的,因为观察到巨变导致许多器官的扩大。尽管如此,研究者还是突出显示了mrps34和spsb3,它们与GWAS目录中的人类身高有关。
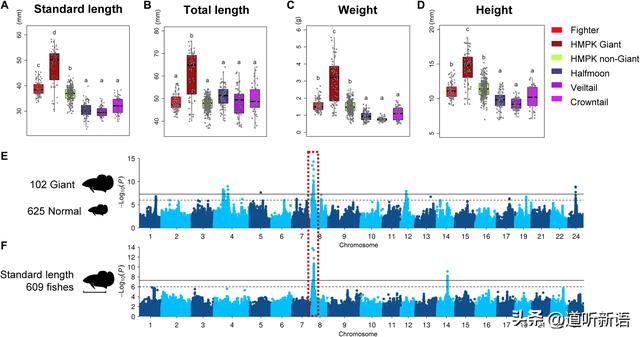
A至D)五组暹罗斗鱼的身体测量。E) 巨型和对照GWAS。 F) 暹罗斗鱼标准体长的GWAS
侵略性
攻击性行为是一种复杂的表型,涉及遗传学,内分泌学,神经生理学和新陈代谢。在一对一的战斗中,暹罗斗鱼的雄性获胜者已被育种者选中,以提高战斗性能,从而产生具有更强侵略性,更长的战斗持续时间和更活跃的战斗型品种。
使用战斗型作为对照,并鉴定了分布在21条染色体上的36个关联峰。S36至S39,表明战斗型与其他品种之间行为差异的多基因基础。在4号染色体上发现最强的关联信号,并且SNP位于两个 esyt2 副本附近。它被证明可以促进果蝇的神经传递和突触生长,有希望成为侵略表型的候选基因。
研究者还强调了由关联峰标记的其他六个神经系统相关基因,包括apbb2编码 β淀粉样蛋白A4前体蛋白结合家族B成员2蛋白,并与神经变性和阿尔茨海默病有关;pank2 编码泛酸激酶,并与神经变性和帕金森综合征有关;crhr2 编码促肾上腺皮质激素释放激素受体2,介导小鼠焦虑;ece2 编码内皮素转换酶-2,调节人类的神经发生和神经元迁移;gpr139 编码孤儿G蛋白偶联受体,是脑回路阿片类药物调节的核心参与者;和 grm3 与双相情感障碍和精神分裂症有关。对这些关联的进一步研究可能会为攻击性行为背后的神经基础提供机械学见解。
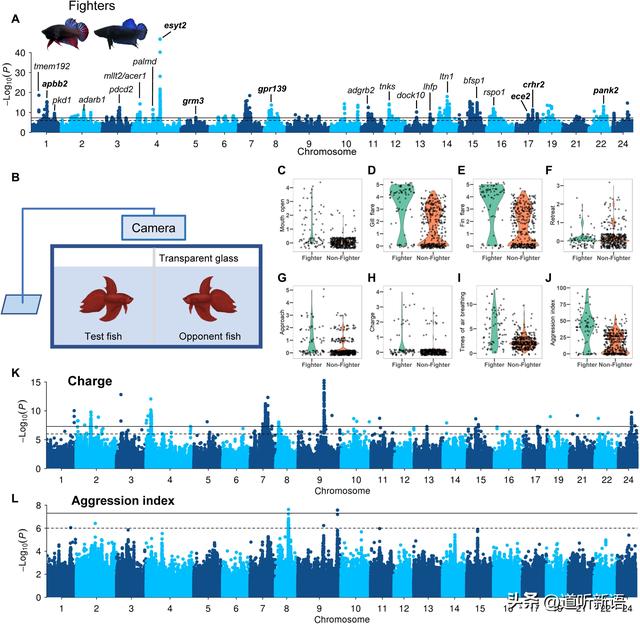
A) 战斗型与对照组 GWAS。 B)量化侵略性方法。 C to J)战斗型与非对照组表型图。 K、L)GWAS分析图。
为了研究与战斗型相关的行为,分析了模拟战斗期间显示的10种不同行为,包括接近,空气呼吸,鳃耀斑,鳍耀斑,抽搐,嘴巴张开,起搏和后退,并进行关联映射以识别每种行为的遗传结构。用于战斗期间行为的GWAS鉴定出一个密切相关的SNP。其位于gfra2的两个拷贝附近,在控制神经元存活和分化中起关键作用。
值得注意的是,关联信号在GWAS中表明战斗期间这两种相关行为的共同遗传基础。通过合成攻击指数中的所有10种行为来量化攻击性,根据该指数,战士型表现出强143%的侵略性。GWAS在侵袭指数上鉴定出8号染色体上的一组相关变异,位于atp5g2基因上或附近,atp5g2基因是线粒体腺苷三磷酸合成酶的亚基。9号染色体上的另一个信号标记了unc-13同源物B基因,这与人类精神分裂症和部分性癫痫的风险有关。
总结
南昌大学洪一江教授团队的结果表明,暹罗斗鱼是了解脊椎动物中许多不同特征的遗传基础的绝佳模型,包括着色和图案,鳍/四肢的骨骼发育以及行为特征,如攻击性。作为易繁殖的动物,暹罗斗鱼作为生物医学研究模型具有巨大的潜力,可以作为现有的脊椎动物模型的补充,如斑马鱼和小鼠。
为确保您能收到每一篇文章,请添加关注啦!
欢迎把我推荐给你的家人和朋友哟~
如果喜欢本篇文章,请点赞、转发和发表您的评论噢~~


















 鲁公网安备37020202000759号
鲁公网安备37020202000759号